|
Politica ecologica - Ecological politics |
|
Giovanni Verzotti -
Home |
Modello di ecosistema.
A life cycle computer model. (1991- additional specifications: 1998 ; 2009)
 According to investigations about ecosystem behaviour by Lotka,Volterra and others, I
propose here a computer simulated matter energized cycle. The matter is
transferred alternatively between a
energy carried population, mainly producers, consumers and decomposers, the
first ones sun power supplied, with intermediate compartments filled by the
same 'dead' population. Living populations differ from dead ones for their
ability, represented by algorithms operating into computer memory,
that return their numeric consistence. The model allows to test balance of population variation, cycle behaviour and
might be used in many cyclic context, such as economy, networks, policy.
According to investigations about ecosystem behaviour by Lotka,Volterra and others, I
propose here a computer simulated matter energized cycle. The matter is
transferred alternatively between a
energy carried population, mainly producers, consumers and decomposers, the
first ones sun power supplied, with intermediate compartments filled by the
same 'dead' population. Living populations differ from dead ones for their
ability, represented by algorithms operating into computer memory,
that return their numeric consistence. The model allows to test balance of population variation, cycle behaviour and
might be used in many cyclic context, such as economy, networks, policy.
A way to describe an ecosystem cycle is:
D x1/D t = f(x n , x 1 , t, e), D xi /D
t = f(x i-1, x i ,
t, e), xi,t
= population living into i compartment at time t, i = 2,3,..n e =
energy, t = time .
It is useful to compute pollution or new temporary
energy sources, but following first and second thermodynamic principles 'wells'
and 'sources' are not allowed (only codes, as DNA, and their expression able to
structure the matter are not conservative). Then any i , any t,
∑ xi,t
= constant ( >0)
Each compartment is a function of the previous one.
Computing with a discrete generation system over an oriented graph, using an
ordinary computer work sheet, I consider three matter compartments (resources
and two “dis-organics” combined only in diagrams), a source of energy
subdivided in renewable and not, producers and its parasites compartment to
exemplify, consumers (zoo) and de-compositors (bacteria). Matter and energy
supplied in discrete bio-quanta ( matter and
energy constituting a cell is a good model of bio-quantum). I mean
“dis-organics” as matter containing energy but not enough information to
"live"; two population (producers and decomposers), operating with
algorithm (information) provided: the first ones (i.e. phototrophs) give energy
charged copy of themselves; the second ones (i.e. bacteria) return resources
during their replica using energy charged matter. In this model populations are
computed with decimals, time flows as a trend of events not as a clock, it is
possible to adjust or change many variables with parameters, algorithms are
function of probability to find 'food' following classic Lotka-Volterra
equations, growth is exponential, it is considered matter only to replicate and
energy to replicate and survive. The model include a sub population of
producers to transform matter, only to duplicate (ratio of these parasitism is
adjustable with parameter); consumers are divided in secondary (zoo) and
primary decomposers ('bacteria'). Secondary consumers must have a distinct
'job' (algorithm or information) otherwise we split
only in two an existing population: here secondary consumers return bulk matter
easier to consume by 'bacteria'. This preference is adjustable with parameter (a ). Growth algorithms are of type:
n t+1= n t
{1+ k p}
where n is
population , p probability to find 'food', k adjustable
coefficient, keeping on account that decomposers (bacteria) return r
resources each replication using energy accumulate in dis-organics. Death
algorithm for each species may be fixed either as percentage of dead elements
over population or related with its enemies, reliability of self devices and
any accident causing irreversible information destruction (here is treated only
percentage and predators probability). It is possible to switch off any
parameter during computation.
Fig. 1- Model used in
computer simulation. Dead are combined in dis-organics only in diagrams
representation.
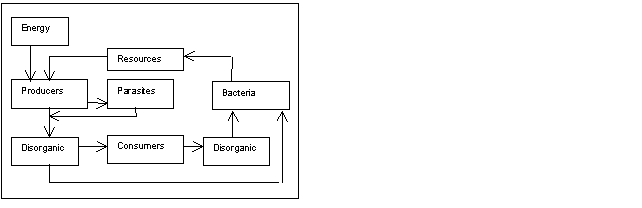
Diagrams
report only the most important variables. For a complete report
refer to program.
Results.
Fig. 2- The simplest situation: resources and
producers (phyto and its parasites at
low rate) with enough energy and without predators. Producers grow reducing
resources and tend to limit r/k (r = resources and k arbitrary ratio of
producer vs. resources)
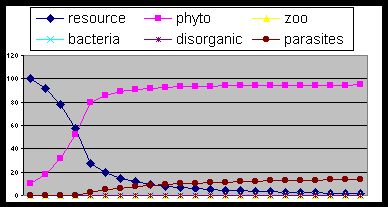
Fig. 3- Introducing decomposers (bacteria) the cycle
is stabilized.
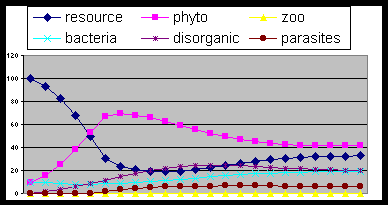
Fig. 4- Adding consumers (zoo) the cycle reach
an equilibrium depending on groups relative ability.
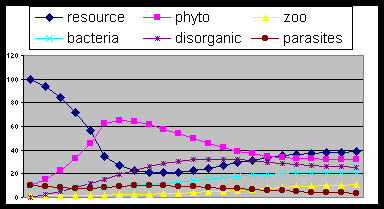
Fig. 5. The same as fig. 3 but with excess of
producers fitness (proa=1). The rest of living
mass is not able to manage the conversion to resources.
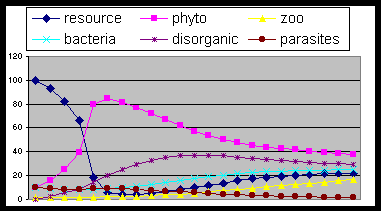
Fig. 6. The same as fig. 3 with excess of zoo
proliferation (propla=1) or phyto destruction: the cycle tends to end owing to
producers plunder.
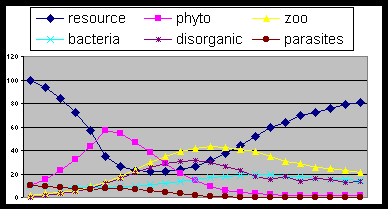
Fig. 7 . The same as fig. 3
with high rate of parasitism (tan = 0,8) that
hampers the cycle progress.
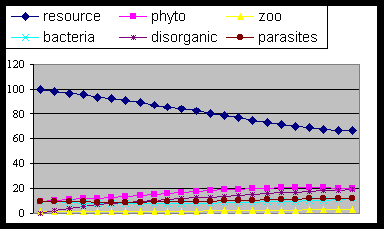
Fig. 8 . The same as fig. 3
with sudden lack of energy at time 11: producers and their parasites die, zoo
and bacteria grow up due to dis-organics increase (organic remains) and the
cycle tends either to end, or to reach the equilibrium allowed by renewable
energy residual, if any. In fact and unfortunately humans have acquired the
ability to change growth and death factors of life cycle, as parameters with
first letter “k..” and “pro..” in the cycle model
listed below.
The least bulk matter remaining (resources and
dis-organics), the best the cycle, keeping on account ties environment. When
environment conditions are defined, any growth exceeding stable steady state is
transitory or clue of drift that may lead to a different steady state .
Consequent remarks
(extracted from letter dated Jan 13th 1993, and published on March
28th 2009).
1) The cycle stops if resource, phototrophs, bacteria
and obviously the sun, tend to lack. We can imagine a mutant man, able to use
CO2, donor and electrons
acceptors, sun energy on his skin, but he would change himself in a vegetable.
2) The matter of all species is the same, changing
only the functions, the organizations and its know
how, then death is necessary. Death sets oneself matter at disposal of others
organisms.
3) Thermodynamics principles are true in spite of
human financial and economics interests.
4) Keeping constant the matter, the more humanity or
new species, the less in other living organisms. The species change itself only
for mutation and each species ought to reach the optimized steady state
(possibly reducing dis-organics at minimum).
5) Only change of mortality and birth rate modify the
system conditions. The species are obliged to production and demolition
functions. Any activity absorbing energy, resources or products changes birth
or death rates. Reciprocally changing birth or death rates change the balance
among species, until extinction with profit of the other .
Should the specie instead learns the function of another one, there is a
compensation among species with exchange or transfer of rolls.
In fact human specie is changing these factors.
6) Increasing or decreasing resources increase populations
but reciprocal ratios don't change.
10) The function and denomination of groups are
determined by their algorithm, that return in a fixed
computer memory location their numeric consistence.
11) In human systems are not important juridical right
but the best informative algorithm related to resources and environment
circumstances, keeping on account that we are intermediate specie, useful but
not necessary in the ecosystem . In case the algorithm doesn't
insure survival, the relevant population (its cultural project) extinguish and
the resources are divided among all the other ones. This is true for each
biological species.
(Note: in economy is nearly the same thing, keeping on
account that human organizations producing objects useful o useless to
surviving with energy consumption, either for their production or for their
use).
Conclusions.
This model of environment behaviour may be improved
but I propose it mainly to redirect political and economical interventions,
provided to critical survival conditions respect: (i)
resources conservation and (ii) matter recycling.
Program list.
The program core in a worksheet cell as follows
(required Framework.II of Ashton Tate
(1986) and perhaps PC with low clock frequency- Full program at
disposal on request ):
@local(indice,zona,avi1,avi2,amo1,mo1,ri1,pla1,plamo1,co1,mo2,ba1,bamo,kbamo,ba,ri,co,mo,pla,avi,ka,kamo,kpla,kplamo,
kco,kb,ktot,tot,alfa,proa,propar,propla,prob,bari,av1,am1,tan,kam1,av0,av2,
source,well,ken,rest),
indice:=1,
zona:=(B6:V27),
tot:=O2*C4+W2*C4+T2*J4+S2+R2*L4+Q2+P2*N4,
ka:=C4,kamo:=D4,kpla:=J4,kplamo:=K4,kb:=N4,kco:=L4,
ba:=P2,ri:=Q2,co:=R2,mo:=S2,pla:=T2,avi:=W2,av1:=O2,
tan:=E4,av0:=O2,kam1:=F4,kbamo:=O4,avi1:=0,amo1:=0, mo1:=0,
ri1:=0,pla1:=0,plamo1:=0, co1:=0,mo2:=0,ba1:=0,bamo:=0,am1:=0,
avi2:=0,av2:=0,v4primo:=0, source :=V4+u3, ken:=V2, ktot:=(kpla+kco),
alfa:=@if(@and(M4<1,M4>0),M4,1),M4:=alfa,
; to display filter on alfa
@while (indice <25, proa:= @if(G4<=0,ri/(tot),G4), ;probability producers find
food for replication
propar:=avi/tot,
; probability parasites find host
ri:= @if(ri>0,ri,0),
@put(zona,ri), ;col.b, resources
@next(zona), ;col.c, producers growth no more than double using ka*resources/prod. + energy /ken
minus cost to pay parasites av0
@if(source>0, ;if1
@if(avi*proa>0,
;if2
@if(source/ken>avi*proa+av0*propar, ;if3 parasites must be rewiewed
owing to growth
@if((ri/ka)>avi*proa+av0*propar, ;if4 depending from avi,but
if av0=0 must not grow
@list(avi1:=avi+avi*proa*(1-tan),
av1:=av0+av0*propar*tan), ;else if4
@list(avi1:=avi+(ri/ka)*proa*(1-tan),
av1:=av0+(ri/ka)*propar*tan) ), ;close if4
else if3
@if((ri/ka)>avi*proa+av0*propar, ;if5
@list(avi1:=avi+source/ken*proa*(1-tan),
av1:=av0+source/ken*propar*tan), ;else if5
@if((ri/ka)>source/ken, ;if6
@list(avi1:=avi+(source/ken)*proa*(1-tan),
av1:=av0+(source/ken)*propar*tan),
;else if6
@list(avi1:=avi+(ri/ka)*proa*(1-tan),
av1:=av0+(ri/ka)*propar*tan)
))), ;close if 6-5-3 else if2
@list(avi1:= avi,
av1:=av0) ), ;close if2, else if1
@list(avi1:=avi,av1:=av0) ), ;close if1
@put(zona,avi1),
@next(zona), ;col.d, producers die at rate kamo
defined as:
kamo:=@if(D4<0,(ba+pla)/tot,D4),;it is possible to fix kamo>0
amo1:=@if(source>0, @if(avi>0,@if(avi1<avi,(avi-avi1)+avi*kamo, avi1*kamo),avi1),avi1),;if
source<=0 all die
@put(zona,amo1),
@next(zona), ;col.e, put parasites computed above
@put(zona,av1),
@next(zona), ;col.f, parasites die at rate kam1
kam1:=@if(F4<0,(ba+pla)/tot,F4),
am1:=@if(source>0,@if(av1>0,
@if(av1<av0,(av0-av)+kam1*av1,av1*kam1),av1),av1),
@put(zona,am1),
av2:=@if(av1-am1>=0,av1-am1,0), ;progr. parasites,change
variab.(for ri)
@next(zona), ;col.g, progr. deaths,converted amo1 e am1 in quanta
mo1:=
amo1*ka+am1*ka+mo,
@put(zona,mo1),
@next(zona), ;col.h progr. producers
avi2:=@if(avi1-amo1>0,avi1-amo1,0),
@put(zona,avi2),
@next(zona), ;col.i progr.resources,protect
from producers reduction ;not parasites because depending from producers
ri1:=ri-@if((avi1-avi)>0,(avi1-avi)*ka+(av1-av0)*ka,0),
@put(zona,ri1), ;avi1 is total
@next(zona),
propla:=@if(H4<=0,mo1/tot,H4),;col j secondary consumers grow eating mo1 no more 2*pla
@if(@and(pla>=0,mo1>=0),
@if(mo1/ktot>pla*propla,
@list(pla1:=pla+pla*propla,
co1:=co+pla*propla,
mo2:=mo1-(pla1-pla)*ktot), ; mo2 in quanta
@list(pla1:=pla+mo1*propla/ktot,
co1:=co+propla*mo1/ktot,
mo2:=mo1-(pla1-pla)*ktot)),
; close if2 now else if1
@list(pla1:=pla,mo2:=mo1,co1:=co) ),
@put (zona,pla1),
@next(zona), ; col k secondary consumers die
kplamo:=@if(K4<0,ba/tot,K4),
plamo1: =@if(pla1>0,pla1*kplamo,pla1),pla:=pla1-plamo1,
;report.in col.r
@put(zona,plamo1),
@next(zona), ;col. l progr. co1 evaluated with pla1
@put(zona,co1),
@next(zona), ;col. m progr mo2, evaluated with pla1,plus plamo1(quanta)
mo2:=mo2+plamo1*kpla,
@put (zona,mo2),
prob:=
@if(I4<=0,(co1*kco+mo2)/(tot),I4),; prob must be
here for ba updating
@next(zona), ;col. n, primary consumers grow eating co1,
then mo2
@if(ba>0, @if(mo2>0,
@if(co1>0, ; if1, if2, if3
@if(kco*co1>(1+kb*prob)*ba, ;if4
@list(co:=co1-prob*ba*(kb+1)/kco, mo:=mo2,ba1:=ba+prob*ba),
@if((kco*co1+mo2)>(1+kb*prob)*ba, ;if5 in else if4
@list(co:=co1*(1-prob),
mo:=mo2-alfa*prob*(ba-co1*kco*prob/(kb+1))*(kb+1),
ba1:=ba+prob*co1*kco/(kb+1)+alfa*prob*(ba-prob*kco*co1/(kb+1))),
; else if5
@list(co:=co1*(1-prob),mo:=(1-alfa*prob)*mo2,
ba1:=ba+prob*(co1*kco/(kb+1)+alfa*mo2/(kb+1))) ;list closed
) ; close if5
), ;if4'closed, else if3=co1<=0
@if(mo2>(1+kb*prob)*ba, @list(co:=co1,mo:=mo2-prob*alfa*ba*(kb+1),
ba1:=ba+alfa*ba*prob),
@list(co:=co1,mo:=mo2*(1-alfa*prob), ba1:=ba+prob*alfa*mo2/(kb+1)))
; close if6
), ; close if3 elseif2 mo2<=0
@if(co1>0,@if(kco*co1>(1+kb)*ba, ;if7 e if8
@list(mo:=mo2,co:=co1-prob*ba*(kb+1)/kco,ba1:=ba+ba*prob),
@list(co:=co1*(1-prob),mo:=mo2,ba1:=ba+prob*co1*kco/(kb+1))
), ;if8 closed else if7= co1<=0
@list(mo:=mo2,co:=co1,ba1:=ba) ) ;if7
), ; now branch else ba<=0 if2
@list(co:=co1,mo:=mo2,ba1:=ba)),
;if1 note:ba are previous ones
@put(zona,ba1),
@next(zona), ;col.o prim. consum
dead (go to ri),protect from dimin. sec. consum.;follow two way to cause mortality put semicolon
before not desired
kbamo:=@if(O4<0,ba1/tot,O4),
bamo:=@if(ba1>0,ba1*Kbamo,ba1),
; bamo:=@if(ba1>0,ba1^2*kbamo/tot,ba1),
bari:=@if((ba1-ba)>0,(ba1-ba),0),
;each plamo eaten 1 resource free
ri:= ri1+bamo*kb+bari, ; (1)**see mo
here cancel bari with option mo not ri
@put(zona,bamo), ;resources in quanta,for every kb
@next(zona), ;col.p progr. sec.
consumers and protect from errors:else gives *FALSE!
ba:=@if(ba1-bamo>=0,ba1-bamo),
@put (zona,ba),
@next(zona), ;col.q,progr resources:sec.
consum.* kb+res. released during growth
@put(zona,ri), ;computed before
ba, because ba updated in
col n
@next(zona), ;col.r, progr calculated with ba1
@put(zona,co), @next(zona), ;col.s, mo the same col. r
; mo:=mo+bari, *(1)**active when sec. consumers become mo,not ri(take away bari in ri)
mo:= mo,
@put(zona,mo), @next(zona), ;col. t,progr
primary consumers pla1-plamo1
@put(zona,pla),
@next(zona), ;col.u,progr. parasites
well:=@if(avi1-avi>0,(avi1-avi)*ken,0),+@if(av1-av0>0,(av1-av0)*ken,0),
;to compute before change of variab.
avi:=avi2,av0:=av2, ;change variables, see col. F
@put(zona,av0),
@next(zona), ;col v
rest:= U3-well,
<CR> v4:=@if(rest>0, V4, V4+rest), source:=u3+v4,
;energy is shared in renewable(u3) added each cycle and
unrenewable(v4)
@put (zona,well),
@next(zona), ; col b next row
indice:=indice+1)
;There is independent matter test, with the same formula for tot (see
program start).